Механізм кросинговеру
За допомогою всієї цієї складної механіки здійснюється тонке вирівнювання гомологічних хромосом по гомології первинної структури ДНК, яке, як ми побачимо нижче, необхідне для кросинговеру.
Внесення розривів і інвазія у більшості еукаріот відбуваються до формування синаптонемного комплексу, забезпечуючи необхідне для нього гомологічне вирівнювання хромосом. Що характерно, невеликі ділянки порушення гомології, наприклад, невеликі інверсії в гетерозиготі (ділянки ДНК, в одному з гомологів повернені на 180о), не перешкоджають формуванню синаптонемного комплексу по всій довжині біваленту. Однак у дрозофіли навпаки, формування синаптонемного комплексу передує розривам. Швидше за все, вона може це собі дозволити тому, що у неї гомологічні хромосоми перебувають у вирівняному стані не тільки в профазі мейозу, але і в інтерфазі.
Коли кількість впізнань досягає якогось критичного рівня, подвійні структури Холідея починають розділятися. Їх розділення відбувається шляхом ліквідації перехресть між двома подвійними ланцюгами ДНК. Це досягається шляхом розрізання двох одноланцюгових ДНК, що створюють перехрестя, і заліковуванні утвореного одноланцюгового розриву.
Структурам Холідея властива так звана ізомеризація. ДНК - достатньо гнучка рухлива молекула, і її обертання в ділянках перехрестя може створювати два варіанти структури Холідея, при якій або нерекомбінантні ланцюги ДНК будуть перехрещеними, а рекомбінантні - неперехрещені, або навпаки, нерекомбінантні ланцюги ДНК будуть неперехрещеними, а рекомбінантні - неперехрещені. На рисунку знак рівності позначає різні площинні представлення однієї і тієї ж ситуації, тобто пов'язані ним картинки топологічно еквівалентні.
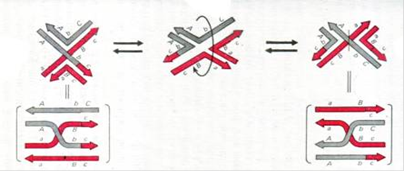
Рис.3.Ізомеризація структури Холідея
Ці два альтернативних стани виявляються далеко не рівноімовірними, і на те, яке саме буде домінувати, впливає присутність низки спеціальних білків.
Розділення структур Холлідея може відбуватися по одному з двох варіантів - або по конверсійному шляху, або по рекомбінаційному шляху. Вибір шляху відповідає одному з ізомерів, показаних вище, але визначається не випадковим вигинанням ниток ДНК, а тим, з якими білками пов'язана та чи інша структура Холідея.
При роздленні по конверсійному шляху структура Холідея розділяється, виходячи з такого ізомеру, де перехрещені ланцюги ДНК, один з яких брав участь в інвазії, а другий - утворював D-петлю. При цьому обидва брали участь у обміні хроматиди (дволанцюгове ДНК) зшиваються так, як вони вступали в обмін, і можуть нести зміни тільки в ділянці, що побувала в складі гетеродуплексів.
Розділення по рекомбінаційному шляху відбувається виходячи з такого ізомеру, де перехрещуються ті ланцюги ДНК, які не брали участь в інвазії і не утворювали петлі. У результаті саме вони виявляються розрізаними в області перехрестя і надалі пришиваються до ланцюгів у складі подвійних ланцюгів, що звільнилися від перехрещення. Як наслідок, обидва ланцюги кожної хроматиди виявляються зшитими з ланцюгами іншої хроматиди, тобто дволанцюгові молекули виявляються зшитими хрест-навхрест. Повторимо ту ж картинку, доповнивши її розділенням структури Холідея, ліворуч - по конверсійному шляху, праворуч - по кросоверному. Нагадаємо, що зображена «ліва» структура і мається на увазі ще одна структура праворуч, причому правіше її два верхні ланцюги ДНК, що прийшли від однієї хроматиди, повністю «сірі», а дві нижні, що прийшли від іншого - «червоні».
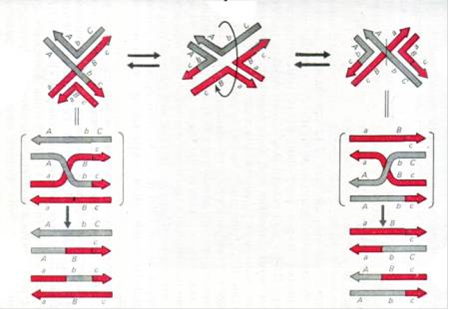
Рис.4.Розділення структури Холідея різними шляхами
Однак ми зараз розглянули одну структуру Холлідея, а у нас їх є дві, з двох боків від проміжної структури. Якщо обидві з них розділяться по конверсійному шляху, то кросинговер не відбудеться. Таку ситуацію легко візуалізувати на вже приведених схемах проміжних структур (повторимо її ще раз, знову в двох крайніх варіантах протяжності міграції гілок у дріжджів і сордаріі), якщо подумки розрізати перехрещені нитки ДНК в місцях перехрещення і зшити їх так, щоб вони не перехрещувалися.
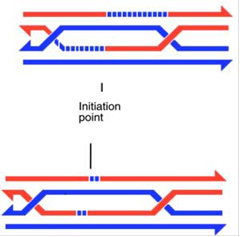
Рис.5.Розділення двох структур Холідея по конверсійному шляху.
Однак при цьому в складі кожного ланцюга в ділянці колишньої проміжної структури або залишаться гетеродуплекси, або вони будуть репаровані по одному з ланцюгів.
Якщо ж хоча б одна з двох структур Холідея розділилася по кросоверному шляху, ми будемо мати кросинговер. На наведеній нижче схемі для проміжної структури дріжджів зліва показаний випадок, коли таким чином розділилася права структура Холідея, праворуч - коли так розділилася ліва структура Холідея.
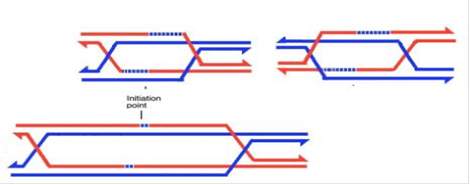
Рис.6.Розділення правої і лівої структур Холідея по кросоверному шляху
Легко побачити, що процедура розрізання перехрестів і зшивання сусідніх кінців призведе тепер до того, що кожен з ланцюгів ДНК кожної з хроматид виявляться пришитими до ланцюгів протилежної хроматиди праворуч і ліворуч від колишньої структури Холідея, тобто ми будемо мати кросинговер. Слід звернути увагу на те, що кросинговер зачіпає обидва ланцюга ДНК задіяних у ньому хроматид, але з деяким зрушенням (на довжину колишньої проміжної структури).
Подібні статті
Ссавці Чернігівської області
Коли після важкої праці виїжджаєш на лоно природи і спостерігаєш, як усе
живе жадібно підставляє себе під сонячне проміння, купається у ньому, коли
дивиться на квітучі луки або обережно підступаєш до таємничого болота, слухаєш
пташиний спі ...
Родина Rosaceae у флорі Бистрицької улоговини (Прикарпаття) та перспективи їх використання
Актуальність роботи.
На сьогоднішній день надзвичайно важливим завданням постає збереження та вивчення біологічної різноманітності на нашій планеті. Проблема збереження біологічної різноманітності як одного із стратегічних завдань людства висвітлен ...