Електронномікроскопічні дослідження апікальних меристем картоплі та вплив хіміотерапії на процес оздоровлення
Досліджували 2 концентрації нових речовин - 0,1 і 0,05 г/л, використовуючи двофазну дію: у першу фазу витримували частинки зелених пагонів картоплі в рідкому розчині антивірусних речовин протягом 2 діб; у другу фазу виділяли експланти розміром 300 - 500 мкм і висаджували їх на поживне середовище з додаванням АВР.
Як контроль під час порівняння ефективності антивірусної дії використовували антивірусний комплекс, який складався із 2,4-діоксогекса-гідро-1.3,5-триазину (ДГТ), ціаногуанідину і розгалуженого манану (РМ) - полісахариду, виділеного із дріжжів роду Candida sp. (клітинні дріжжові манани).
Проводили комплексне тестування отриманих ліній рослин-регенерантів, використовуючи методи імуноферментного аналізу, електронної мікроскопії і рослин-індикаторів.
Електронномікроскопічні дослідження ультратонких зрізів меристемних тканин картоплі дають певне уявлення про розвиток вірусної інфекції в них, а також можливість виявити деякі деталі механізму розвитку інфекції.
При вивченні меристем досліджували такі питання: характер будови та ультраструктурну організацію клітин меристематичної тканини картоплі по зонах; наявність плазмодесм в клітинах на різних рівнях; розподіл і спосіб локалізації вірусних включень у меристематичних клітинах.
Деталі ультраструктурної організації меристемних клітин були встановлені на прикладі сорту Приєкульська рання. В результаті проведених досліджень серій ультратонких зрізів встановлено, що меристемні тканини в зоні 10-15 мкм від верхівки складаються з клітин без вакуолей, цитоплазма має рівномірний гомогенний матрикс із слабо диференційованими клітинними органелами. Починаючи з зони 20-30 мкм вже спостерігається диференціація клітинних органоїдів. З'являються вакуолі, матрикс ядра більш просвітлений. Характерним для цих ділянок тканин є наявність у вакуолях більшості периферійних клітин порівняно великих глобулярних тіл, можливо, саме там накопичуються в клітині рістактивуючі речовини (рис. 2). Ядерця в клітинах, як правило, мали центральні просвітлені зони, які не спостерігалися в ядерцях звичайної листової тканини.
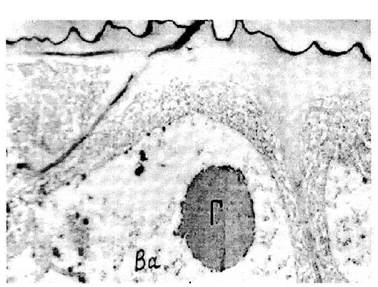
Рис. 2. Глобулярні включення у вакуолях меристематичних клітин. Зона розміром близько 30 мкм від верхівки. Ва - вакуоль, Г - глобули. Збільшення -19000.
Особливий інтерес для розуміння механізм)' проникнення вірусів у клітини меристемних тканин викликає процес формування плазмодесм, які є своєрідними каналами розповсюдження вірусів у звичайних рослинних тканинах. Відсутністю плазмодесм окремі дослідники пояснюють відсутність вірусу в меристемній тканині [9].
Дослідження показали, що перші плазмодесми, ще не зовсім сформовані, виявляються вже в зоні близько 30 мкм, добре сформовані плазмодесми виявляли в зоні близько 60 мкм.
Із всіх досліджених вірусів досить легко виявляли МВК в меристематичних тканинах, починаючи із зони 80-90 мкм. Локалізація вірусних включень є доволі різною: загальною тенденцією є те, що віруси зустрічаються здебільшого біля ядра. Форма і розмір вірусних включень теж різні, від невеликих включень, до масових. Потрібно відмітити здатність М-вірусу утворювати кристалічні структури по типу паракристалів (рис.3). Як правило, такий спосіб локалізації в клітині характерний для жорсткопа-личкоподібних вірусів і, зокрема, для вірусу тютюнової мозаїки.
Подібні статті
Ряд вугреподібні (Anguilliformes)
Вугреподібні - загадка яку люди
намагались розгадати вже більше 2 тисячі років. Майже всі вугреподібні, 22
родини з приблизно 350 видами, - морські риби, що живуть переважно в теплих
морях, але представлені й на більших глибинах. Тільки од ...
Ссавці Чернігівської області
Коли після важкої праці виїжджаєш на лоно природи і спостерігаєш, як усе
живе жадібно підставляє себе під сонячне проміння, купається у ньому, коли
дивиться на квітучі луки або обережно підступаєш до таємничого болота, слухаєш
пташиний спі ...