Взаємодія фагів з бактеріями
Отже, в результаті дії вегетативного фага у зараженій бактерії з'являється значна кількість нових корпускул фагів і ми говоримо про репродукцію фагів бактеріальною клітиною на основі генетичної інформації, заданої нуклеїновою кислотою батьківського фага. Саме в цьому й виявляється своєрідна форма паразитизму фагів на субклітинному молекулярному рівні.
Внутрішньоклітинний розвиток у фагів, які містять різні типи нуклеїнової кислоти, дещо відрізняється за характером її реплікації, зокрема, одноланцюгова ДНК і РНК фага спочатку повинні набути дволанцюгової реплікативної форми, а вже після цього в клітині нагромаджуються нові молекули відповідної фагової нуклеїнової кислоти.
Водночас із формуванням дозрілих вібріонів у бактеріальній клітині утворюються літичні ферменти, детерміновані нуклеїновою кислотою фага. Ці ферменти можуть розкладати цупкий пептидо-глікановий шар клітинної стінки; з їхньою допомогою здійснюється четверта стадія взаємодії фага з бактеріальною клітиною — лізис клітинної стінки і вихід нового потомства бактеріофагів назовні (рис. 9).
Літичний, або продуктивний, цикл розвитку характерний для вірулентних фагів, які є справжніми паразитами бактерій. Однак у природі поширеними є й так звані помірні фаги. При зараженні ними бактерій гине тільки невелика частина клітин, а решта нормально розмножується і стає носіями відповідних помірних або симбіотичних фагів. Явище фагоносіння бактеріями дістало назву лізогенії.
Докладне вивчення показало, що існують псевдолізогенні та справжньолізогенні бактеріальні культури. Переважна більшість клітин першого типу є стійкою до цього фага і тільки невеличка кількість їх може заражатися фагом і давати його репродукцію. Справжньолізогенні — це теорія несе в собі фаг у певній прихованій формі і може за відповідних умов репродукувати його.
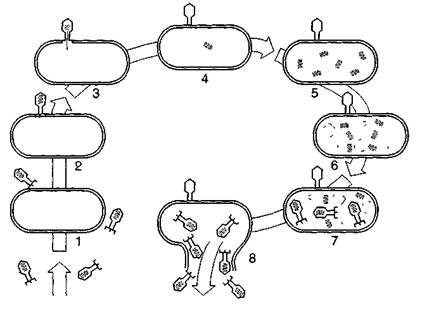
Рис. 9. Схема циклу розмноження бактеріофага Т2:
1 — фаги оточили бактерію, 2 — вібріон фага прикріплюється до клітини, 3 — в клітину впорскується вірусна ДНК, 4 — капсид фага залишається ззовні бактерії, 5 — синтезуються нові молекули ДНК, 6 — утворюються білкові оболонки фагів, 7 — відбувається збирання нових вірюнів, S— бактерія руйнується (лізується), І вібріони фага виходять назовні.
Встановлено, що особлива форма фага, яка перебуває у справжньо-лізигенних бактеріях (профаг) є нуклеїновою кислотою (геном фага), яка тісно інтегрована з генетичним матеріалом бактеріальної клітини, і в разі поділу бактерії передається її потомству. Отже, в лізигенній клітині профаг поводить себе як нормальний її компонент.
Важливою властивістю лізигенної культури є її стійкість до фагів, які містяться в ній. У зв'язку з цим вивчення помірних фагів лізигенної культури можливе тільки тоді, коли є інша культура цього виду, чутлива до помірного фага даної лізигенної культури. Такі культури дістали назву індикаторних.
Лізогенія дуже поширена серед усіх систематичних груп мікробів. Вона спостерігається у збудників черевного тифу і паратифу, дифтерійної палички, спороносних і бульбочкових бактерій, дріжджів, пеніцилу тощо.
Профаг лізигенної культури може спонтанно або в разі індукції перетворитися на дозрілий бактеріофаг. Натомість у деяких випадках під впливом різних чинників у профага виникають мутації, в результаті яких при індукції він не здатний перетворюватися на повноцінну фагову частинку. Внаслідок цього в середовище можуть виділятися дефектні фаги, що складаються тільки з однієї головки або відростка. Такі фаги можуть адсорбуватися на бактеріях, але не можуть розмножуватися у них. Дефектні фаги привернули до себе увагу вчених, оскільки, як виявилось, багато описаних бактеріцинів є дефектними фагами. Дефектна лізогенія дуже поширена в природі.
Подібні статті
Паразитичні пласкі черв'яки
Проблеми забезпечення здоров'я населення країни є сферою
міжгалузевого регулювання і одним з найважливіших аспектів соціальної політики
в області охорони здоров'я населення [Демьянков Е. Н., 1996].
Сучасний рівень розвитку медицини вимаг ...
Зимові скупчення сороки в Любечі
Біологія та чисельність зимуючих птахів залишається недостатньо вивченою
не лише в Україні, але й у більшості європейських країн. Це зумовлене
труднощами проведення польових досліджень у цей період, відсутністю
уніфікованих методик, а тако ...