Закономірності інтерференції
У дріжджів обидва варіанти розділення структур Холідея приблизно рівноймовірні. У багатоклітинних велика частина D-петель розділяється за конверсійним шляхом, і лише деякі - за рекомбінаційним.
У дріжджів розділення по рекомбінаційному вимагає присутності білка MSH4, до якого потім приєднуються білки MLH1 і MLH3. У ссавців відбуваються множинні посадки MSH4 на хромосоми в момент їх спаровування, їх кількість все зменшується, і нарешті в тих сайтах, де з'являються білки MLH1 і MLH3, відбувається кросинговер. Відсутність будь-якого з двох білків виключає у ссавців більш 90% кросоверних подій. Інші 10% кросинговеру йдуть по якомусь іншому механізму, з залученням інших білків.
Всі згадані білки належать до родини білків mismatch repair і беруть участь у репарації гетеродуплексів. І всі вони безпосередньо взаємодіють один з одним під час кросинговеру.
На стадії пахітени в місцях, в яких відбувся кросинговер, під електронним мікроскопом видно досить великі, близько 90 нм, білкові гранули - пізні рекомбінаційні вузлики, що представляють собою комплекси вищезазначених і, можливо, якихось ще білків. Як ми вже говорили, флюоресцентні антитіла до MLH1 також добре маркують точки майбутнього кросинговеру вже на стадії пахітени, як це візуалізувано на фотографії П.М. Бородіна (місця посадки MLH1 тут дрібні точки, що світяться зеленим, тоді як великі - це центромери):

Рис.7. Точки майбутнього кросингговеру на стадії пахітени
На цій стадії хромосоми компактизовані ще не дуже сильно і, головне, рівномірно, тому відстань між сусідніми сайтами посадки MLH1 більш-менш адекватна фізичній відстані (уздовж молекули ДНК). Ця відстань варіює, і щоб зрозуміти як саме, слід побудувати і проаналізувати його статистичний розподіл.
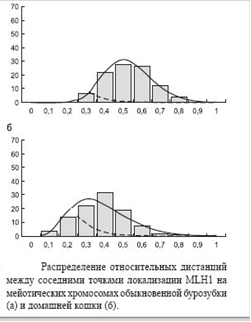
Рис.8. Розподіл відносних дистанцій між сусідніми точками локалізації MLH1
Розподіл Пуассона кількості випадкових подій на відрізку шкали довжиною T
P (n) = хn e-х / n!
(де x = λT) не є єдиним розподілом, який характеризує простий пуассонівський процес. Можна розглянути також розподіл довжини інтервалів шкали між сусідніми випадковими подіями. Воно буде описуватися так званим показовим розподілом.
P (y) = λ e-λy
Зауважимо, що якщо розподіл Пуассона задано на множині невід'ємних цілих чисел, що показове - на безлічі невід'ємних дійсних чисел. Всупереч інтуїтивним очікуванням, цей розподіл має максимум в нулі і J-подібну форму (тобто воно монотонно зменшується в міру збільшення y).
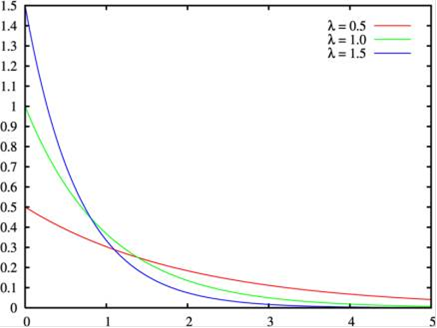
Рис.9.Розподіл Пуассона
Як нам тепер і слід було очікувати на підставі існування інтерференції, реальний розподіл відстаней між точками посадки MLH1 не відповідає показовому розподілу. Перш за все, воно має максимум, відмінний від нуля. Однак воно виявилося в дивовижній відповідності до гамма-розподілу, вірніше розподілу Ерланга. Гамма-розподіл має додатковий параметр k. Якщо він заданий на множині натуральних чисел, то воно називається розподілом Ерланга. Це розподіл також породжується простим пуассоновським процесом. Воно є розподіл інтервалів шкали не між сусідніми, а між k-ми сусідніми подіями: від кожної події до другого наступного за k = 2, до третього при k = 3 і т. д. Показовий розподіл - це розподіл Ерланга з k = 1 . На жаль, розподіл Ерланга не висловити простими формулами. Він при великих k наближається до нормального.
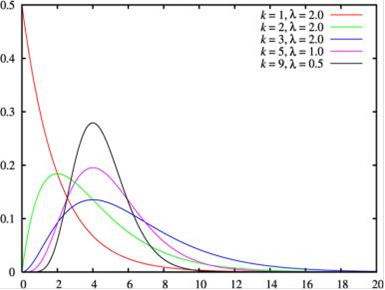
Рис.10. розподіл Ерланга
Виходить, що в основі кросинговеру все ж лежить простий пуассонівський процес, але він властивий не кросинговеру, а передуючій йому події, тоді як кросинговер відбувається на кожному k-му такій події. Передуючою подією тут є дволанцюговий розрив, який розвивається в проміжну структуру. Відстань між ними якраз таки повинна відповідати розподілу Пуассона, однак візуалізувати їх не вдається, оскільки розриви відбуваються на стадії лептотени, коли хромосоми ще занадто довгі і переплутані.
Проміжні структури розділяються по конверсійному або кросоверному шляху. Виходить, що існує якийсь механізм, що змушує розділятися по кросоверному шляху кожну k-у проміжну структуру. Природа цього механізму поки невідома. Не виключено, що якісь білки маркують проміжні структури і при цьому збираються в агрегати - k-(або близько того), залишаючи k +1 структуру вільною для зборки комплексу білків, необхідних для розділення структури Холідея по кросоверному шляху.
Подібні статті
Медоносні рослини луків
Прекрасний і багатогранний рослинний світ, і хоч пройшло багато тисячоліть з часу його виникнення, непізнаність і нерозгаданість його вабить дослідників. Якби ми знали, скільки втрачаємо, коли не робимо жодних спроб пізнати те, що нас оточує, що дає ...
Молекулярні механізми кросинговеру
Кросинговер - це явище обміну ділянками
гомологічних хромосом після кон’югації у профазі-1 мейозу. Результатом цього
процесу є обмін генетичною інформацією між хромосомами. Це є одним з головних
чинників еволюції,так як забезпечує комбінат ...