Закономірності інтерференції
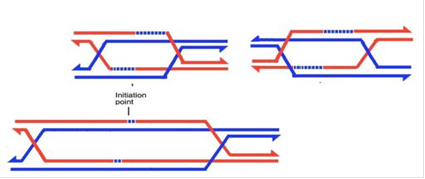
Рис.6.Розділення правої і лівої структур Холідея по кросоверному шляху
Отже, беремо дріжджі (зверху), подумки розрізаємо перехрещені ланцюги, знаходимо примикаючі до перехрестя гетеродуплекси і репаруємо їх по нерозрізаному ланцюгу. Зліва маємо тільки один гетеродуплекс у верхній хроматиді, який репаруюється по червоному ланцюгу при розділенні по конверсійному шляху, в результаті чого отримуємо октаду 4: 4 без конверсії, і по синьому ланцюгу при розділенні по кросоверному шляху, з конверсією 6: 2. Праворуч гетеродуплекс розміщений знизу, а результат протилежний: він буде репарований по червоному ланцюгу при розділенні по кросоверному шляху, з результатом 4: 4, та по синьому ланцюгу при розділенні по конверсійному шляху, з результатом 6: 2. Отримуємо, що в половині випадків кросинговер не буде супроводжуватися конверсією, а в половині - буде супроводжуватися конверсією 6:2.
У сордаріі (знизу), внаслідок протяжної міграції гілок краю проміжної структури являють собою симетричні протяжні ділянки гетеродуплексів. Репарація при розділенні структур Холлідея дає тетради 4:4 без конверсії, проте велика частина гетеродуплексів залишаються не порушеними такою репарацією і дають аберантні 4: 4 октади, тобто октади з постмейотичною сегрегацією.
На завершення, враховуючи усі згадані вище можливості, ми можемо розписати повну групу можливих подій у наступній схемі, де колонки по чотири рядки позначають чотири хроматиди, з яких верхня і нижня не взаємодіяли, а дві середні формували проміжні структури. Тут A, a і B, b позначають алелі крайових генетичних маркерів, + і m позначають домінантний або мутантний алель, за яким ми стежимо в аналізі; середні два символи в кожному короткому рядку, розділені косою рискою позначають алелі локусу m в подвійних ланцюгах ДНК, тобто гомо-або гетеродуплекси.
Конверсії нема Конверсія
•Норм. 4:4 абер. 4:4 5:3 6:2
• Кросинговеру нема
•A m/m B A m/m B A m/m B A m/m B
•A m/m B A m/+ B A m/m B A m/m B
•a +/+ b a +/m b a m/+ b a m/m b
•a +/+ b a +/+ b a +/+ b a +/+ b
• Кросинговер є
•A m/m B A m/m B A m/m B A m/m B
•A m/m b A m/+ b A m/m b A m/m b
•a +/+ B a +/m B a m/+ B a m/m B
•a +/+ b a +/+ b a +/+ b a +/+ b
Загадковий другий шлях кросинговеру
У аскоміцетів повного збігу розподілу кросоверів з рахунковою моделлю кроссинговера не спостерігаються. Досить складні дані про конверсії і рекомбінації свідчать, що у них є два шляхи кросинговеру.
. «Ранній» шлях: msh4-незалежний кросинговер без інтерференції (вірніше зі слабкою негативною інтерференцією), який повністю визначається подіями в момент інвазії однонитого кінця і, можливо, не пов'язаний з утворенням D-петлі і знайомої нам проміжної структури, тобто в чомусь схожий з моделлю Холідея. Які саме при цьому утворюються проміжні структури та яким чином вони вирішуються - поки невідомо. У дріжджів, у яких репарація гетеродуплексів в ділянці інвазії майже облігатна, вона в нормі супроводжується конверсією поблизу точки інвазії за типом 6:2. У сордаріі, у якої репарація тетеродуплексів в районі інвазії відбувається далеко не завжди, він супроводжується конверсією за типом 5:3, рідше 6:2.
. «Пізній» шлях: msh4-залежний кросинговер з позитивною інтерференцією у відповідності зі лічильної моделлю, який відбувається під час розділення структур Холідея. У дріжджів він в половині випадків пов'язаний з конверсією типу 6:2, у сордаріі - з відсутністю конверсії поблизу точки, де сталося розділення структури Холідея по кроссоверному шляху.
З цим були пов'язані отримані на сордарії результати Кітанова, опубліковані в 1978, який зробив наступний висновок: є два типи кросинговеру:
Кросинговер з інтерференцією, який не супроводжується конверсією
Кросинговер без інтерференції, який супроводжується конверсією.
Дійсно, конверсія поблизу точки ініціації обміну спостерігається пов'язаною з подіями раннього неінтерфереруючого кросовера,. У той же час репарація гетеродуплексів поблизу структур Холідея, яка відбувається в момент їхнього розділення за рахунок пізнього інтерферуючого кросинговеру, робить конверсію невидимою у сордаріі, тоді як конверсія поблизу точки ініціації при довгих проміжних структурах сордаріі відбувається досить далеко і не асоціюється з подією кросинговеру.
Зауважимо, що вивчати зв'язок конверсії та кроссинговера можна лише маючи багато відповідних генетичних маркерів, що допускає тетрадний аналіз, і порахувавши астрономічні кількості тетрад. Проте абсолютно надійні і багатозначні результати, отримані Кітановим в 1974 р., по-перше, протягом чотирьох років залишалися неопублікованими, оскільки суперечили результатам, отриманим на дріжджах, які казали, що будь-який кросинговер супроводжується конверсією поблизу від точки свого проходження (ми вже знаємо чому ), і її можна зареєструвати, якщо там є відповідний ген. По-друге, ці результати на тридцять років опинилися на узбіччі генетики, не отримавши ні підтвердження, ні пояснення, ні розвитку. Після закриття його лабораторії в Йокогамі в 1989 р., Кітанов віддався вивченню китайської і англійської поезії і садівництву. І тільки в 2008 році група під керівництвом Френкліна Сталя - того самого, який експериментально встановив напівконсервативний характер реплікації ДНК - повернулася до робіт Кітанова і досліджувала те ж питання на дріжджях, підтвердивши наявність другого типу кросинговеру, якому не властива інтерференція.
Подібні статті
Психрофільні мікроорганізми та їх використання
В даному рефераті мова
піде про психрофільні мікроорганізми, що широко представлені в тваринному світі.
До них належать представники про – та еукаріотів, а також багато видів грибів, дріжджів
та навіть комах і хребетних. В останні роки впл ...
Динаміка та еколого-біологічні особливості дендрофлори пам’ятки місцевого значення саду імені Т.Г Шевченко м. Харкова
Однією з характерних особливостей сучасної епохи є невтримний темп урбанізації. Створюючи і розвиваючи міста, людство змінює вигляд навколишнього середовища. Місто, як штучна формація, різко відрізняється від своєї географічної зони. Змінюються мікро ...