Закономірності інтерференції
В 70-х роках минулого століття у дріжджів спостерігався зв'язок між конверсією і кросинговером ,він передбачав, що вони є різними проявами одного й того ж процесу. Ці спостереження істотно просунули розуміння молекулярного механізму кросинговеру, але з'ясований він все ж був досить недавно.
Ми зараз цілком можемо дати пояснення генної конверсії, виходячи зі знання механізму кросинговеру, точніше його початкових етапів. Ми не дарма називали один із шляхів розділення структур Холлідея конверсійним. Коли обидві структури Холлідея однієї проміжної структури розділяються по цьому шляху, цілісність вихідних хроматид відновлюється, але в тому місці, де вона перебувала, залишаються ділянки гетеродуплексів. Подивимося ще раз на рис.5. Розділення двох структур Холідея по конверсійному шляху (тут червоним і синім позначені ланцюги ДНК спочатку однієї несестринскої хроматиди , а пунктиром - ДНК, добудована по матриці відповідного кольору).
Згадаймо ще, що за кадром у нас залишилися дві хроматиди, що не беруть участь в обміні. В ділянці, де відбувалося добудовування другого ланцюга ДНК ми бачимо асиметричне розташування гетеродуплексу: так що в цій ділянці три ланцюги ДНК одного кольору і один - іншого кольору. Додаючи по два ланцюги кожного кольору від необмінних хроматид ми отримуємо розщеплення 5: 3. Якщо гетеродуплекс випадково захопив варіабельний сайт (ділянка, по якому розрізняються алелі) досліджуваного нами локусу, то ми будемо спостерігати октади (продукти одного мітозу, наступного за одним мейозом) саме з таким співвідношенням алелів, як, наприклад, на ось цій фотограціі:
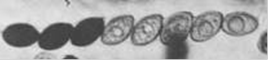
Рис.11.Октади
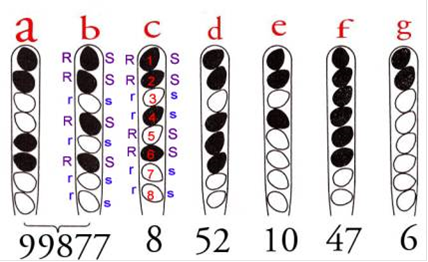
Рис.12.Тетради
В ділянці, порушеної міграцією гілок, де добудовування не відбувалося, ми спостерігаємо симетричне розташування гетеродуплексів, яке не порушує співвідношення ниток ДНК різного кольору, так що ми будемо мати співвідношення ланцюгів ДНК 4: 4. Хоча співвідношення ниток і не порушено, ми можемо розпізнати отримані октади як аберантні, оскільки внаслідок постмейотичної сегрегеціі порушене звичне нам розташування аскоспор з ідентичним алелем суміжними двійками, як наприклад у випадку з попередньої картинки.
Питання - звідки беруться конверсійні тетради з співвідношеннями 6: 2, як у випадках f і g вище і на наступних фотографіях?

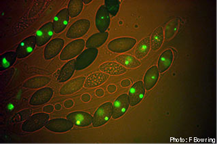
Рис.13,14. Конверсійні тетради з співвідношеннями 6: 2
Розглянуті нами механізми не дають відповіді на це питання, оскільки такі тетради з'являються в результаті ще одного, додаткового процесу. Цей процес - мейотична репарація гетеродуплексів (meiotic mismatch repair). Репарація гетеродуплексів ініціюється однонитчастим розривом і виправляє надрізаний ланцюг по цілому. У нашій системі вона може відбуватися в двох місцях, де утворюються дуплекси з однонитчастих розривів - в ділянці інвазії одночасно з інвазією і біля структур Холідея в момент їхнього розділення.
Біля точки інвазії репарація гетеродуплексів незмінно має місце у одних об'єктів і з невеликою ймовірністю відбувається у інших. Якщо вона відбувається з невеликою ймовірністю, як у сордарії, то переважно виникають конверсійні октади 5:3. Якщо вона має місце завжди, як у дріжджів, то репарація ланцюга, що мав надріз, по матриці нерозрізаного ланцюга ДНК призводить до уніфікації в цьому місці всіх чотирьох ланцюгів ДНК двох хроматид, що беруть участь в проміжній структурі. З урахуванням ланцюгів ДНК двох хроматид, що не беруть в ній участі, - ми отримаємо співвідношення ланцюгів ДНК 6:2. Це можна вивести зі схеми 5 Розділення двох структур Холідея по конверсійному шляху, звернувши увагу, що в ділянці такої проміжної структури репарація гетеродуплексів по матриці нерозрізаних ланцюгів ДНК зробить всі чотири ланцюги ДНК «синіми»:
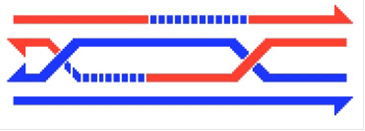
Рис.
Репарація гетеродуплексів при розділенні структур Холідея і біля них як мінімум у сумчастих грибів йде завжди. При цьому вона ініціюється однонитчастими розривами ДНК, в даному випадку виникнені при розрізанні перехрещених ланцюгів, а матрицею служать неперехрещені ланцюги. Зрозуміти її результати нам допоможе повторення наведеної вище схеми варіантів розділення структур Холідея, де для однієї напівхіазми показаний конверсійний шлях, а для іншої - кросоверний. Ми зараз переконаємося, що в силу асиметрії проміжної структури у дріжджів репарація гетеродуплексів при розділенні «правої» і «лівої» напівхіазми приведуть до різних результатів, незалежно від того, по якому шляху вона розділилась:
Подібні статті
Концепція еволюції в біології
Поняття еволюції вживається в різних змістах, але здебільшого ототожнюється з розвитком. Усі чули про глобальну еволюцію Всесвіту, геологічну еволюцію й еволюцію живої природи. У всіх цих випадках під еволюцією мався на увазі процес тривалих, поступо ...
Хвороби та шкідники квіткових рослин
Протягом багатьох століть пояснення хвороби були примітивними. Розвиток їх почався з ХІХ століття після доведення хибності теорії про зародження організмів.
Основоположники лісової фітопатології Антуан де Барі, Воронін.
Фітопатологія – вивчає ...